| VIVO Патофизиология | Эндокринная система > | Механизмы действия гормонов |
Механизм действия: гормоны с рецепторами клеточной поверхности
Белковые и пептидные гормоны, катехоламины, такие как адреналин, и эйкозаноиды, такие как простагландины, находят свои рецепторы, украшающие плазматическую мембрану клеток-мишеней.
Связывание гормона с рецептором инициирует серию событий, которые приводят к генерации так называемых вторичных мессенджеров внутри клетки (гормон является первым мессенджером). Затем вторые посланники запускают серию молекулярных взаимодействий, которые изменяют физиологическое состояние клетки. Другой термин, используемый для описания всего этого процесса, – это преобразование сигнала .
Структура рецепторов клеточной поверхности
Рецепторы клеточной поверхности являются интегральными мембранными белками и, как таковые, имеют области, которые участвуют в трёх основных доменах:
- Внеклеточные домены: некоторые из остатков, находящихся вне клетки, взаимодействуют с гормоном и связывают его – другим термином для этих областей является лиганд-связывающий домен .
- Трансмембранные домены: гидрофобные участки аминокислот «удобны» в липидном бислое и служат для закрепления рецептора в мембране.
- Цитоплазматические или внутриклеточные домены: хвосты или петли рецептора, которые находятся в цитоплазме, реагируют на связывание гормона, каким-то образом взаимодействуя с другими молекулами, что приводит к образованию вторичных мессенджеров. Таким образом, цитоплазматические остатки рецептора являются эффекторной областью молекулы.
Было идентифицировано несколько отличительных вариаций в структуре рецептора. Как показано ниже, некоторые рецепторы представляют собой простые однопроходные белки; многие рецепторы факторов роста принимают эту форму. Другие, такие как рецептор инсулина, содержат более одной субъединицы. Другой класс, который включает бета-адренорецепторы, проходит через мембрану семь раз.
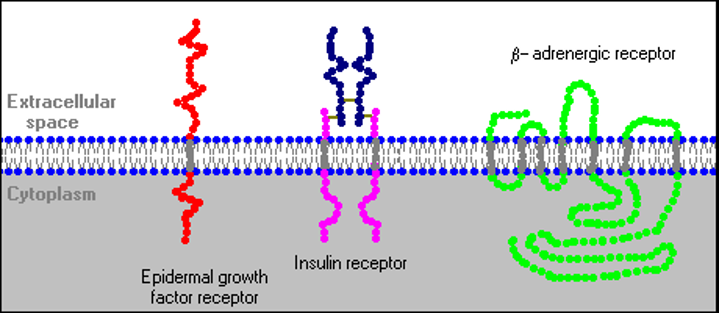
Молекулы рецепторов не изолированы сами по себе и не фиксируются в одном месте плазматической мембраны. В некоторых случаях другие интегральные мембранные белки взаимодействуют с рецептором, чтобы модулировать его активность. Некоторые типы рецепторов группируются вместе в мембране после связывания гормона. Наконец, как подробно описано ниже, взаимодействие гормонально-связанного рецептора с другими мембранными или цитоплазматическими белками является ключом к генерации вторичных мессенджеров и передаче гормонального сигнала.
Системы второго мессенджера
Подумайте, что бы произошло, если бы поздно ночью вы заметили горящее здание. Надеюсь, что вы наберёте 911 или аналогичный номер службы экстренной помощи. Вы должны сообщить диспетчеру о пожаре, а диспетчер, в свою очередь, свяжется с несколькими пожарными и «активирует» их. Затем пожарные сразу же приступили к работе, поливая огонь водой, устанавливая контрольно-пропускные пункты и тому подобное. Они также, вероятно, активируют других «игроков», таких как полиция и следователи по пожарной безопасности, которые придут позже, чтобы попытаться определить причину пожара. Важно отметить, что как только пожар потушен (или здание полностью разрушено), пожарные возвращаются на станцию и ложатся спать.
Реакция сообщества на пожар, по крайней мере в некотором смысле, аналогична системе вторичных посыльных, участвующей в действии гормона. В описанном сценарии вы являетесь «первым посланником», диспетчер – «приёмником», пожарные – «вторыми посланниками».
В настоящее время в ячейках распознаются четыре системы вторичного обмена сообщениями, как показано в таблице ниже. Обратите внимание, что не только несколько гормонов используют одну и ту же систему вторичных мессенджеров, но и один гормон может использовать более одной системы. Понимание того, как клетки интегрируют сигналы нескольких гормонов в согласованный биологический ответ, остаётся сложной задачей.
| Второй посланник | Примеры гормонов, которые используют эту систему |
| Циклический AMP | Адреналин и норадреналин , глюкагон , лютеинизирующий гормон, фолликулостимулирующий гормон , тиреотропный гормон, кальцитонин , паратиреоидный гормон , антидиуретический гормон |
| Активность протеинкиназы | Инсулин , гормон роста , пролактин , окситоцин , эритропоэтин , несколько факторов роста |
| Кальций и / или фосфоинозитиды | Адреналин и норадреналин , ангиотензин II , антидиуретический гормон , гонадотропин-рилизинг-гормон, тироид-рилизинг-гормон. |
| Циклический GMP | Натуретический гормон предсердий , оксид азота |
Во всех случаях, казалось бы, слабый сигнал, генерируемый связыванием гормона с его рецептором, усиливается внутри клетки, превращаясь в каскад действий, которые изменяют физиологическое состояние клетки. Ниже представлены два примера систем вторичного мессенджера, обычно используемых гормонами. В качестве примеров используются глюкагон и инсулин, оба из которых в конечном итоге работают через молекулярный переключатель, включающий фосфорилирование белка. Имейте в виду, что в обоих случаях очень сложная система значительно упрощается.
Системы второго мессенджера Cyclic AMP
Циклический аденозинмонофосфат (цАМФ) представляет собой нуклеотид, образующийся из АТФ под действием фермента аденилатциклазы . Внутриклеточная концентрация цАМФ увеличивается или уменьшается за счёт различных гормонов, и такие колебания влияют на множество клеточных процессов. Один известный и важный эффект повышенных концентраций цАМФ является активация цАМФ-зависимой протеинкиназы называется протеинкиназы А .
Протеинкиназа А номинально находится в каталитически неактивном состоянии, но становится активной, когда связывает цАМФ. После активации протеинкиназа А фосфорилирует ряд других белков, многие из которых сами являются ферментами, которые либо активируются, либо подавляются путём фосфорилирования. Такие изменения ферментативной активности внутри клетки явно изменяют её состояние.
Теперь давайте соберём эту информацию, чтобы понять механизм действия гормона, такого как глюкагон :
- Глюкагон связывает свой рецептор на плазматической мембране клеток-мишеней (например, гепатоцитов).
- Связанный рецептор взаимодействует с набором G-белков и включает аденилатциклазу, которая также является интегральным мембранным белком.
- Активированная аденилатциклаза начинает преобразовывать АТФ в циклический АМФ, что приводит к повышению внутриклеточной концентрации цАМФ.
- Высокие уровни цАМФ в цитозоле делают вероятным, что протеинкиназа А будет связываться с цАМФ и, следовательно, каталитически активна.
- Активная протеинкиназа A «бегает по клетке», добавляя фосфаты к другим ферментам, тем самым изменяя их конформацию и модулируя их каталитическую активность – – – абра-кадабра, клетка изменилась!
- Уровни цАМФ снижаются из-за разрушения цАМФ-фосфодиэстеразой и инактивации аденилатциклазы.
В приведённом выше примере действие гормона заключалось в изменении активности ранее существовавших компонентов в клетке. Повышение цАМФ также оказывает важное влияние на транскрипцию определённых генов.
Системы второго мессенджера тирозинкиназы
Рецепторы некоторых белковых гормонов сами по себе являются протеинкиназами, которые активируются связыванием гормона. Киназная активность, связанная с такими рецепторами, приводит к фосфорилированию остатков тирозина на других белках. Инсулин является примером гормона, рецептором которого является тирозинкиназа.
Гормон связывается с доменами, выставленными на поверхности клетки, что приводит к конформационному изменению, которое активирует киназные домены, расположенные в цитоплазматических областях рецептора. Во многих случаях рецептор фосфорилирует себя как часть процесса активации киназы. Активированный рецептор фосфорилирует множество внутриклеточных мишеней, многие из которых являются ферментами, которые активируются или инактивируются при фосфорилировании.
| Рисунок справа предназначен для изображения рецептора тирозинкиназы, подобного тому, который используется в инсулине. После связывания гормона рецептор претерпевает конформационные изменения, фосфорилирует себя, а затем фосфорилирует множество внутриклеточных мишеней. | 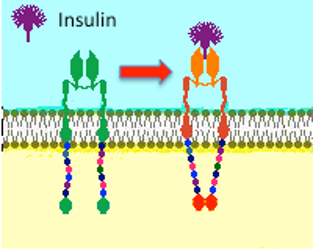 |
Как было замечено с системами вторичного мессенджера цАМФ, активация рецепторных тирозинкиназ приводит к быстрой модуляции ряда белков-мишеней внутри клетки. Интересно, что некоторые из мишеней рецепторных киназ представляют собой протеинфосфатазы, которые после активации рецепторной тирозинкиназой становятся компетентными для удаления фосфатов из других белков и изменения их активности. Опять же, кажущееся небольшое изменение из-за связывания гормонов усиливается во множестве эффектов внутри клетки.
В некоторых случаях связывание гормона с поверхностным рецептором вызывает каскад тирозинкиназы, даже если рецептор сам по себе не является тирозинкиназой. Рецептор гормона роста является одним из примеров такой системы – взаимодействие гормона роста с его рецептором приводит к активации цитоплазматических тирозинкиназ с результатами, концептуально аналогичными результатам, наблюдаемым в случае рецепторных киназ.
Судьба гормонально-рецепторного комплекса
Нормальное функционирование клетки зависит от того, являются ли каскады вторичных мессенджеров временными событиями. Действительно, ряд видов рака связан с рецепторами, которые постоянно стимулируют системы вторичных мессенджеров. Одной из важных составляющих негативного регулирования действия гормонов является интернализация рецепторов клеточной поверхности. Во многих случаях интернализация стимулируется связыванием гормонов.
Интернализация происходит за счёт эндоцитоза через структуры, называемые покрытыми ямками. Образовавшиеся эндосомы (иногда называемые «рецептосомами») могут сливаться с лизосомами, что приводит к разрушению рецептора и гормона. В других случаях оказывается, что гормон диссоциирует и рецептор повторно используется путём слияния эндосомы обратно в плазматическую мембрану.
| Гормоны с внутриклеточными рецепторами |
Отправляйте комментарии на Richard.Bowen@colostate.edu